
Wen-Sung Chung of the University of Queensland takes us on a deep dive into the complex and contradictory world of cephalopod vision and colour use. How are colour-blind animals able to display such a flamboyant variety of colours? Read on to find out
This blog is part of our colourful countdown to the holiday season where we’re celebrating the diversity and beauty of the natural world.
Click here to read the rest of the colour countdown series

Coleoid cephalopods (cuttlefish, squid and octopus, BUT not nautilus) are a group of fast-moving molluscs with the camera-like eyes and the highly centralised nervous system which are rarely seen in invertebrates. In common with their major competitors, the fish, these visually-dominant coleoids display a variety of adaptations with respect to optical properties such as (1) a graded-refraction spherical lens to form a focused image in the aquatic realm; (2) independent eye movements conducted with lens accommodation to extend visual field; (3) dynamic screening pigment migration to adjust light sensitivity; (4) underlying neural circuits to boost sensitivity for dim environments. These adaptations are often linked to the light conditions across a great range of depths in the ocean from well-lit reefs to constantly dark deep waters, resulting in a famous case of convergent evolution in eyes between two animal groups.
Most would agree coleoids are more versatile in rapid and dynamic colouration (<1 second) than a flatfish (seconds or minutes). These soft-bodied coleoids, particularly those dwelling in coastal waters, are the masters in their ability to alter their visual appearance from camouflage, emotional expression, to intra- and inter-specific communication throughout dynamic changes of body patterns. It is particularly impressive in performing aposematic patterns to emphasise their existence or startle potential threats (e.g. blue-ringed octopus and flamboyant cuttlefish) as well as a sudden switch between matching the tone and texture to different backgrounds (see two examples of algal octopus below). Also, they are renowned in rapid visual learning to solve sophisticated tasks (e.g. shape and size discrimination, catching a crab in a jar and collaborative-hunting with fish) and gain experience by observation or through conditioning training. The colourful and dynamic body patterns of coleoids and their ability to match backgrounds for camouflage raise the fundamental question of whether they have colour vision like many fishes? Do they see what fishes see? How do they produce variable body patterns?

The biological display panel – cephalopod skin
Many studies on the behavioural neurobiology of coleoids reveal that their remarkably complex centralise nervous system is in charge of visually-guided reactions to novel challenges such as ballistic tentacular strike, observational learning. Most famous cases in this respect are the remarkable camouflage, mimicry and other communication abilities using dynamic body patterns. The coleoid skin comprises of a system of the neurally-driven elastic coloured chromatophores and the underlying passive reflective layers of leucophores and iridophores, forming a biological display panel parallel to a television screen. In general, the chromatophores are controlled by a set of brain lobes where visual cues are integrated to generate display patterns (from a simple colour spot, chromatic pulse to rhythmic passing waves). Apart from chromatic components, coleoid skin patterns can be further enhanced by a textural component (papillae), especially benthic octopus and cuttlefish. Coleoid camouflage utilises a combination of colouration and papillae, thus making other predators which possess colour vision struggle to spot them in the natural habitat where their body shape is perfectly blending in the surroundings.
Colour-blind camouflage

Although coleoids can display complex colour patterns, anatomical, physiological and behavioural studies indicate that most are colour-blind. Firstly, in order to discriminate colours, this requires the comparison of at least two visual pigments of different spectral sensitivity such as many fishes possessing at least dichromatic (e.g. blue and green sensitive receptors) or trichromatic (e.g. blue, green and red receptors) colour vision. For instance, the trigger fish can learn to pass an Ishihara-style colour discrimination test. In contrast, examinations of retina amongst coastal cephalopod species demonstrate that these shallow water dwellers apparently possess a single photoreceptor type with a single type of blue-green sensitive visual pigment, rendering their visual world entirely monochromatic (further reading, see Chung & Marshall 2016, Proceedings B. doi:10.1098/rspb.2016.1346). It is in some ways similar to the rod-dominant retina of nocturnal or deep-sea fish which is adaptive for luminance and unlikely for seeing colour.
Along with the simple and uniform photoreceptors with a single visual pigment, behavioural evidence in coastal coleoids provides compelling proof of colour-blindness. Behavioural approaches found that the European common octopus, Octopus vulgaris, was capable of recognising objects based on brightness levels, but failed in all colour discrimination trials. Similar results were found in common European cuttlefish, Sepia officinalis, which when tested were unable to match their body colour to that of the gravel background (e.g. red gravels on white background or yellow on blue gravels). Instead, cuttlefish showed different body patterns to match the background using intensity instead of wavelength cues. As many coastal coleoids live in well-lit environments and are under intense predatory pressure, it is perhaps surprising that they have not evolved colour vision for predatory avoidance, mating interactions or indeed their own also very aggressive predatory feeding style. Ironically, humans still struggle to spot cephalopods in the natural habitat where their colour-blind camouflage is perfectly matched with the surroundings.
Alternative visual cue – polarisation signals
Although cephalopods seem to ignore colour information from the environment, they are able to see polarised light, another property of light which is invisible to human eyes (Further reading, see Temple et al 2012, Current Biology doi: 10.1016/j.cub.2012.01.010 and Temple et al 2021, J Exp Biol doi: 10.1242/jeb.240812). The polarisation of light is often treated as something special and unusual; however, along with colour (e.g. visible wavelength for human eyes ca. 400-700 nm) and intensity (brightness), polarisation is the third physical property that describes how the electric field of the light moves through space. Polarised light in the aquatic environment provides an alternative visual cue to those who can detect it (e.g. crustaceans and cephalopods). Recently, behavioural approaches demonstrated that the reef cuttlefish and octopus have notably sensitive polarisation vision to detect polarisation contrast. In other words, they see polarisation much as other animals see colour, potentially for enhancement of object detection.
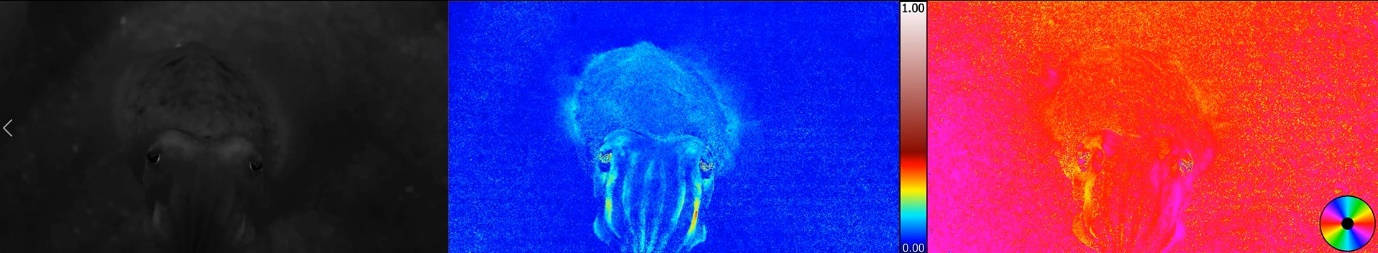
Notably, our current knowledge about how cephalopods might use polarisation sensitivity in their daily life remains sparse. For instance, in situ behavioural observations demonstrate that the day reef octopus can undertake a long foraging path over a large area in one trip but take a short-cut way back to the same den. This is reminiscent of insect (e.g. ant, bee and dung-beetle) navigational ability using polarised light from the sky, but as yet no controlled evidence of this ability has been found in octopus. Another unique feature is that strong polarised signals are seen on arms and other body parts in both squid and cuttlefish due to the high-reflectance iridophores and these polarisation patterns have been proposed to have an inter- or intra-specific communication function. As the cephalopods were among the first animals to evolve complex visual abilities, it is fascinating to speculate that polarisation vision may have evolved before colour vision and indeed in shallow water environments it has some advantages.
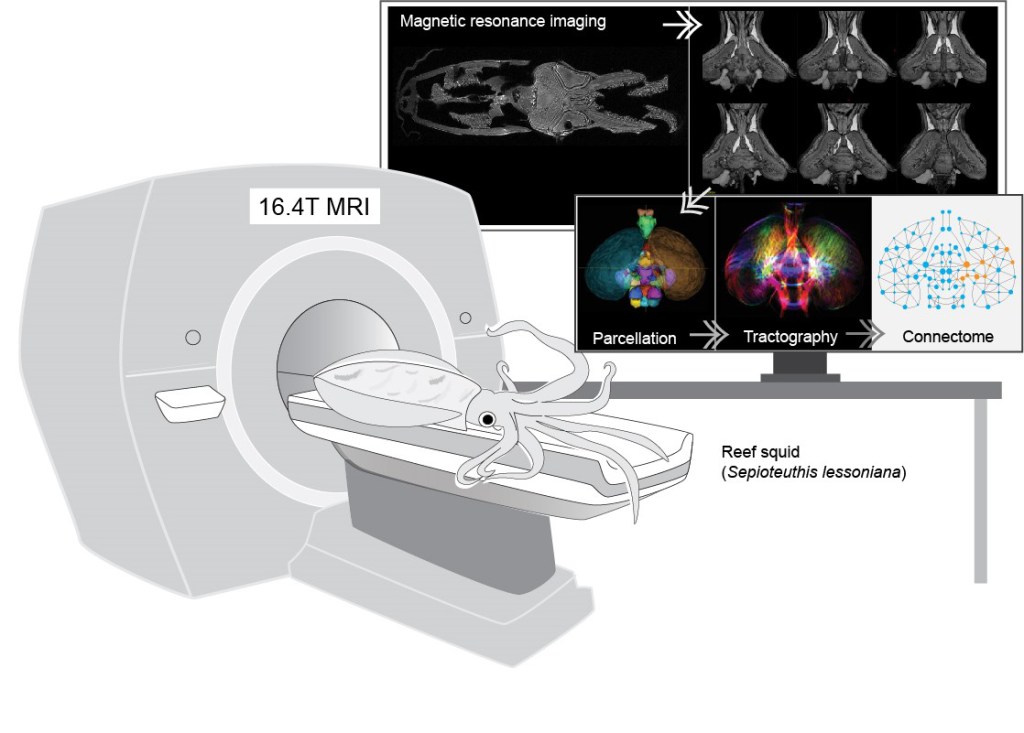
In an effort to understand how the coleoid polarisation vision works, we are currently using the custom made polarisation vision underwater camera (inspired by the cephalopod eye) to simulate vision through the eyes of animals and looking for advantages of this visual information during interactions between conspecifics as well as in their natural habitat. Furthermore, we have successfully adopted the modern day brain imaging technique, magnetic resonance imaging (MRI), to investigate the cephalopod central nervous system in order to further understand how they process visual input throughout the multi-layered visual motor control area and then aim to test whether the hypotheses mentioned above could held. These ongoing approaches aim to provide new insights into the complexity and visual neuroscience of these super-charged mollusc and form a firm grounding to prompt a non-anthropomorphic interpretation of their comparative cognitive and behavioural abilities.
Read the rest of our Colour Countdown series here
Further reading
- Chung, W.-S., and Marshall, N.J. (2016). Comparative visual ecology of cephalopods from different habitats. Proc Biol Sci 283, 20161346. doi:10.1098/rspb.2016.1346
- Chung, W.-S., Kurniawan, N.D., and Marshall, N.J. (2020). Toward an MRI-based mesoscale connectome of the squid brain. iScience 23, 100816. doi: 10.1016/j.isci.2019.100816
- Temple, S.E., Pignatelli, V., Cook, T., How, M.J., Chiou, T.H., Roberts, N.W., and Marshall, N.J. (2012). High-resolution polarisation vision in a cuttlefish. Curr Biol 22, R121-122. doi: 10.1016/j.cub.2012.01.010
- Temple, S.E., How, M.J., Powell, S.B., Gruev, V., Marshall, N.J., and Roberts, N.W. (2021). Thresholds of polarization vision in octopus. J Exp Biol, jeb.240812. doi: 10.1242/jeb.240812

4 thoughts on “Can cephalopods see what fishes see?”